- 首页 > 非靶向植物代谢组学
非靶向植物代谢组学
以植物为研究对象,研究不同物种、不同基因类型或不同生态类型的植物在不同生长时期或受某种刺激前后的小分子代谢产物。基于液质联用技术(LC-MS),无偏向性、尽可能多地检测植物样本内所有的小分子代谢物,对实验组和对照组进行对比分析,通过统计分析筛选差异代谢物,对差异代谢物进行代谢通路分析,进而寻找代谢物与生理病理变化的相对关系。
华大高分辨植物代谢数据库(BGI HRAM-PMDB 1.0)

技术优势
1. 高质量自建库
所有代谢物均通过标准品建库,均包含一级质谱、二级质谱和保留时间信息
2. 代谢物鉴定准确度高
60种标准品校正保留时间,100% 比对标准品鉴定,鉴定数可达1500+
3. 规范化平台严格质控
全流程标准操作程序(SOP)指导,同位素内标与QC双重质控
4. 超高分辨率质谱平台
使用Orbitrap系列谱仪进行检测,仪器分辨率高、质量精度高、稳定性好
5.分析创新
创新性引入相关性差异分析,通过多维信息整合,稳健算法直击核心功能,精准锁定关键机制。
产品应用
1. 植物生长发育研究
2. 农作物品质性状与育种改良研究
3. 植物生物和非生物逆境互作研究
4. 果蔬花卉营养和色泽代谢研究
5. 药用植物功能与活性成分研究
6. 基因功能解析与代谢通路研究
技术路线

技术参数
1. 仪器平台
LC-MS/MS:液相-Waters ACQUITY UPLC,质谱-Thermo Q Exactive / Q Exactive HF / Q Exactive HF-X
2. 分析软件
Compound Discoverer(Thermo, USA),metaX (华大自主开发的代谢组学分析软件包)
3. 数据库
华大高分辨植物代谢组数据库( BGI HRAM-PMDB),包含保留时间、一级质谱和二级质谱信息,化合物数量2500+
mzCloud 在线标准品数据库,包含一级质谱和二级质谱信息,化合物数量30,000+
项目周期
30-60个自然日
案例一:代谢组学研究植物-线虫互作机制[1]
研究背景:
根际微生物及线虫能很大程度地影响植物的健康,其中代谢物小分子信号转导在植物根际的相互作用发挥着重要作用。通常植物和动物在受到病原体攻击时,可以通过识别病原体相关的进化保守的大分子(PAMPs)来诱导激活免疫反应,这种免疫激活的方式被称为pattern-triggered immunity(PTI)。线虫蛔苷是植物寄生线虫分泌的一种保守的糖脂,可以通过PTI诱导植物免疫反应,在植物根际相互作用中起着重要作用,但是目前线虫蛔苷诱导植物免疫的分子机理还未被发现。

图1 植物通过识别和代谢ascr#18介导免疫反应
技术路线:
通过对不同植物(拟南芥、番茄、小麦)在不同处理的条件下(假单胞菌DC3000侵染、线虫侵染、外援物质添加)的组织及根系分泌物进行代谢组分析,发现植物可以通过代谢植物寄生线虫分泌的线虫蛔苷ascr#18生成一系列蛔苷混合物来介导免疫反应。
主要结论:
1. 对比是否经过ascr#18处理的植物组织的代谢数据,发现植物能将ascr#18代谢成一系列蛔苷的混合物如ascr#10,ascr#1,ascr#9,其中ascr#9是含量最丰富的蛔苷代谢物;
2. 通过比较野生型及突变型拟南芥(β氧化关键酶缺失菌株)经ascr#18处理后的组织代谢物水平,观察出植物可以通过β氧化将ascr#18代谢成其他蛔苷;
3. 分别对ascr#18处理后的野生型及突变型拟南芥做假单胞菌DC3000及线虫侵染实验,发现ascr#18代谢是提高植物对线虫侵染的抗性所必需的,但不是植物抵抗细菌所必需的;
4. 对野生型及突变型拟南芥进行ascr#18处理后,发现ascr#18可能不是通过诱导防御基因的表达来提高植物对线虫的抗性的
5. 野生型拟南芥根系能够分泌ascr#18的代谢物ascr#10,ascr#1及ascr#9,其中ascr#9与ascr#18相结合可以抑制线虫对植物的侵染。
案例二:代谢组学结合转录组学探究高修饰脂肪酸的合成途径[2]
研究背景:
虽然初级脂肪酸可以抵抗病原体,但是植物通常还会对这些脂质加以修饰用于应对生物胁迫。其中炔类脂肪酸这种修饰的氧化脂质广泛分布于各种植物中,用于抵抗病原体,但其合成和代谢途径却并没有被发现,这限制了我们对植物抗性的认知及对这类脂质的开发利用。

图2 代谢组学结合转录组学探究法卡林二醇的合成途径
技术路线:
采用非靶向代谢组学及转录组测序相结合的方法,对不同微生物处理后的番茄样本进行分析,筛选出与番茄抗性有关的代谢物及其相关的基因,确定并验证与代谢物相关的基因簇,并进一步对其进行功能探究。
主要结论:
1. 在不同微生物胁迫的番茄样本中,包括法卡林二醇在内的一系列代谢产物的表达发生了明显改变。
2. 通过将转录组和代谢组数据关联分析,发现6个与法卡林二醇表达量更相关的基因,其中4个位于12号染色体的同一区域组成基因簇,可能与其代谢有关。

图3 法卡林二醇的EIC及其6个候选乙炔酶基因在不同样本中的表达量关联图
3. 结合瞬时表达及代谢组实验对基因簇的相关酶功能进行的验证发现两个乙炔酶、一个去饱和酶及一个暂未表现出活性的蛋白;基因突变试验及代谢实验的结果则该基因簇是证明法卡林二醇的合成所必需的。
4. 番茄野生型植株和突变株与病原菌的相互作用试验则表明法卡林二醇的合成基因簇参与法卡林二醇诱导的番茄免疫反应,基因簇上相关基因的缺失也会导致番茄抗性的改变。
参考文献
[1] Manohar, Murli, et al. "Plant metabolism of nematode pheromones mediates plant-nematode interactions." Nature Communications 11.1 (2020): 1-11.
[2] Jeon, Ju Eun, et al. "A Pathogen-Responsive Gene Cluster for Highly Modified Fatty Acids in Tomato." Cell 180.1 (2020): 176-187.
1、数据预处理及质控分析
使用Compound Discoverer软件进行LC-MS/MS数据处理,主要包括峰提取、峰对齐和代谢物鉴定等一系列分析。Compound Discoverer导出的数据通过metaX进行数据预处理、质控分析和后续分析。数据质控的内容包括,QC样本的BPC重叠图、所有样本的PCA分析、各组的CV分析。

图1 数据质控分析
2、 代谢物的检测和鉴定情况
(1)代谢物检测数和鉴定数统计
对数据预处理后的代谢物检测数量和鉴定数量统计如下表。
表1 代谢物检测数和鉴定数统计表
|
离子模式 |
代谢物检测数 |
代谢物鉴定数 |
|
pos |
14604 |
1058 |
|
neg |
12484 |
579 |
pos—正离子模式,neg—负离子模式。
(2)代谢物的分类注释和功能注释分析

图2 代谢物的分类注释和功能注释统计图
3、差异代谢物的筛选
通过PCA分析观察了解两组样本的差异情况。采用多变量分析PLS-DA模型前两个主成分的VIP值,结合单变量分析差异变化倍数(Fold Change, FC))和T检验(Student's t-test)的q-value值来筛选差异表达的代谢物。筛选条件: 1)VIP ≥ 1;2) Fold-Change ≥1.2 或者 ≤ 0.8333;3) q-value < 0.05,同时满足这三个条件,即为差异代谢物。

图3 差异代谢物的筛选
4、差异代谢物分析
包括差异代谢物的聚类热图分析和差异代谢物的代谢通路富集分析。

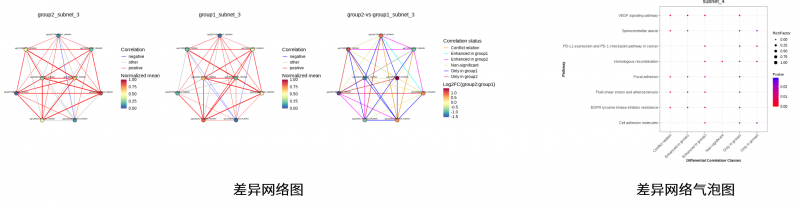
图4 差异代谢物的分析
1、 重复数要求
每组样本的生物学重复次数要求如下,重复次数越多越好。
|
样本类型 |
建议重复次数 |
|
植物样本 |
≥ 6 个 |
2、 送样量要求
样品类型 | 建议送样量/例 |
新鲜植物组织 | ≥500 mg |
冻干植物组织 | ≥ 200 mg |
中药药汤 | 冻干粉≥200 mg,药液≥1mL |
中药药丸 | ≥1g |
Q1:“非靶向植物代谢组学”的代谢物鉴定数量是多少?
A1:代谢物鉴定数(“RT+MS+MS2”或“MS+MS2”鉴定)一般可以达到800-1500,鉴定结果的准确度和数量在行业内属于领先水平。代谢物鉴定数的多少也与样本本身的含有的代谢物数量多少有关,不同物种、不同部位的样本代谢物数量不同,如果样本中的代谢物数量本身就比较少,那么检测和鉴定到的代谢物数量也可能会少一些,以具体项目的检测鉴定情况为准。
Q2:华大“非靶向植物代谢组学”的代谢物鉴定结果可信度如何?
A2:华大“非靶向植物代谢组学”代谢物鉴定使用“RT+MS+MS2”或“MS+MS2”与标准品数据库进行比对,具有很高的可信度。